![Genetics [10] reproduction 3: Meiosis 2](https://apbiology.cn/wp-content/uploads/sites/8/2013/11/genetics-9-dinosuar.jpg)
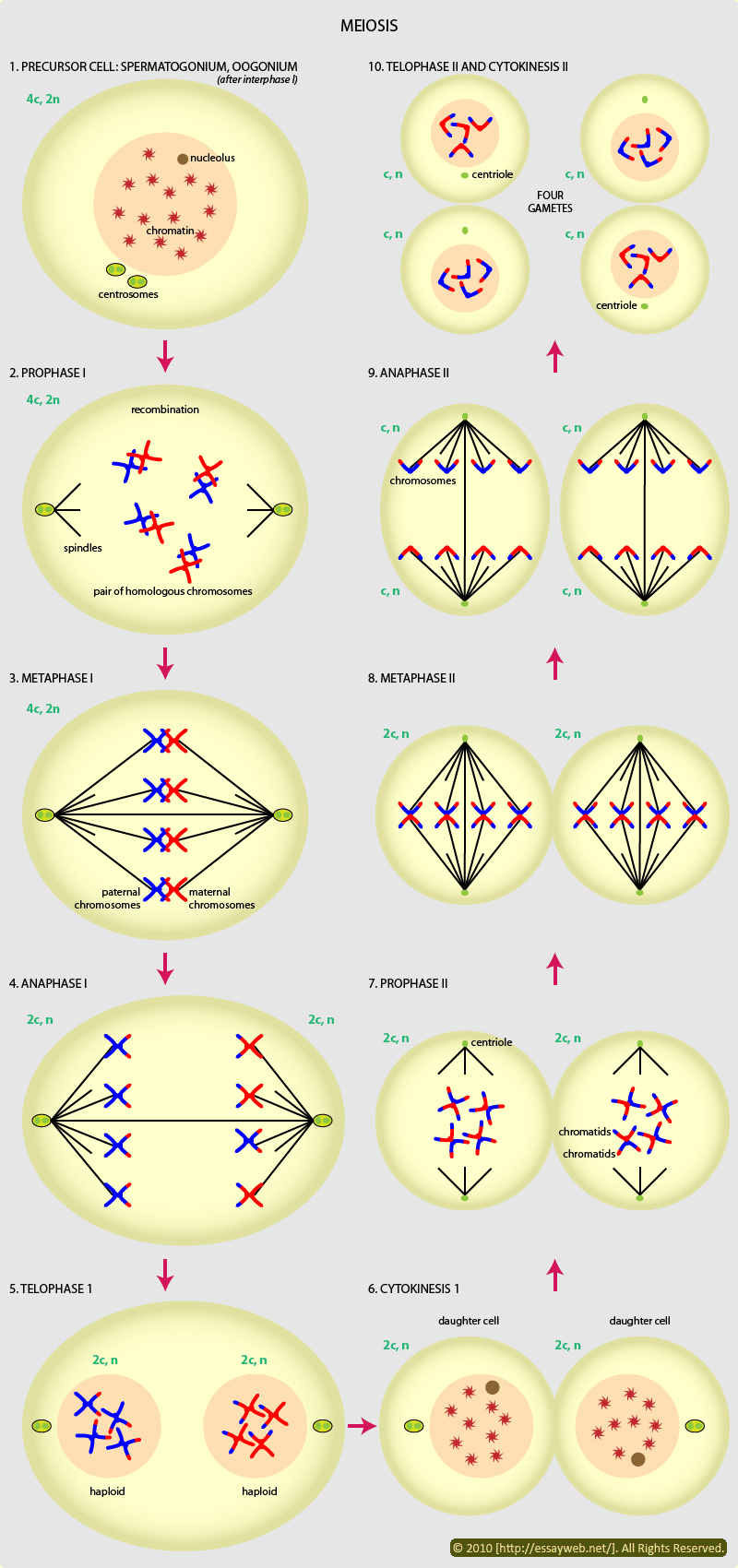
Last section we gave a general introduction of meiosis in terms of chromosome behaviors.
In this section, we will deal with two other respects of meiosis. First, a specific description of meiosis in animals; second, we will mention a situation when meiosis goes wrong.
Meiosis in animals
is found only in ovaries (卵巢)and testes(曲细精管), and even in these tissues is restricted to cells that are destined to form gametes(the germline).
Despite the fact that the mechanisms of gametogenesis differ somewhat between organisms,the steps involved in gametogenesis in mammals are relatively similar.
- In male gametogenesis (spermgenesis).
precursors of germ cells go through many rounds of mitotic divisions in order to maintain a pool of spermatogonia(plural of spermatogonium 精原细胞).↓
Spermatogonia subsequently differentiate into primary spermatocytes(based on what you learned in high school. guess what this word mean? : D ). It is in these cells that meiosis takes place.↓
After meiosis I, these cells are referred to as secondary spermatocytes. These are haploid. The products of the second meiotic division are spermatids. ↓
Spermatids differentiate into motile spermatozoa with rounded or elongate head and a long posterior flagellum.(The final activation of spermatozoa takes place after copulation/sexual intercourse)
Without cytoplasm and many subcelluar organelles, the sperm is light and fast (almost all its weight concentrated in its head, where stored the key of a potential life, nuclei. ).
- In female mammals, the pattern of oogenesis is superficially similar.
Here oogonia(plural of oogonium, 卵原细胞) go through mitotic divisions before differentiating into primary oocytes. These then undergo meiosis.↓
Both daughter cells of the primary oocytes are haploid but differ greatly in size. The larger daughter cell is the secondary oozyte, the smaller the first polar body. The two cells remain attached. Both undergo a second meiotic division.↓
The secondary oocyte undergoes an unequal division producing a large ovum and a small secondary polar body. The first polar body divides into two secondary polar bodies.
Only the ovum, which contains almost all the cytoplasm, will transmit genes into the next generation.
The first meiotic division is only completed at ovulation(the discharge of a mature ovum from the ovary), and the second occurs after fertilization.
Little known: In human females, primary oocytes can be held in meiotic arrest for up to 45 years. This may be important in the increased frequency of aneuploid births observed in older mothers.
Production of aneuploid gametes
The major cause of anueploidy(the situation of having or being a chromosome number that is not an exact multiple of the usually haploid number) is aberrant chromosome behavior at meiosis. In other words, it’s the failure of chromosmes to segregate properly(known as nondisjunction)
At anaphase I, if two homologous chromosomes move to the same pole, first division nondisjunction occurs. In this case, of the 4 cells arsing from meiosis, two will be disomic(contain two copies of the chromosome) and two nullisomic (contain no copy of the chromosome.
At anaphase II, if the chromatids in a cell remain together, division nondisjunction occurs. The resulting tetrad will contain two normal cells, one nullisomic, and one disomic.
Aneuploidy also arises due to nondisjunction at an early mitosis in the embryo, resulting in two populations of cytogenetically different cells in the individual, which is known as a mosaic.
Mosaic is common in Turners syndrome.
↑Turner syndrome or Ullrich–Turner syndrome is a chromosomal abnormality in which all or part of one of the sex chromosomes is absent or has other abnormalities . In some cases, the chromosome is missing in some cells but not others, a condition referred to as mosaicism or “Turner mosaicism”.
Occurring in 1 in 2000– 1 in 5000 phenotypic females, the syndrome manifests itself in a number of ways. There are characteristic physical abnormalities which affect many but not all people with Turner syndrome, such as short stature,swelling, broad chest, low hairline, low-set ears, and webbed necks. Girls with Turner syndrome typically experience gonadal dysfunction (non-working ovaries), which results in amenorrhea (absence of menstrual cycle) and sterility. Concurrent health concerns may also be present, including congenital heart disease, hypothyroidism (reduced hormone secretion by the thyroid), diabetes, vision problems, hearing concerns, and many autoimmune diseases.Finally, a specific pattern of cognitive deficits is often observed, with particular difficulties in visuospatial, mathematical, and memory areas.
Turner syndrome is named after Henry Turner, the endocrinologist who first described it in 1938.(wikipedia)
![Genetics [9] Reproduction 2: Meiosis](https://apbiology.cn/wp-content/uploads/sites/8/2013/11/genetics10-meiosis.gif)
![Mendel’s Genetics [7]: handling problems](https://apbiology.cn/wp-content/uploads/sites/8/2013/12/chi-square.jpg)
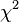 test)=
sum[(obseved expected)ˆ2/expected]
test)=
sum[(obseved expected)ˆ2/expected]
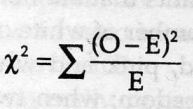
![Mendel’s Genetics [6]: Examples of epistasis](https://apbiology.cn/wp-content/uploads/sites/8/2013/12/genetics-3-snapdragons.jpg)
![Mendel’s Genetics [5]: The dihybrid cross](https://apbiology.cn/wp-content/uploads/sites/8/2013/07/genetics2.png)
![Mendel’s Genetics [4]: examples of mutiple alleles](https://apbiology.cn/wp-content/uploads/sites/8/2013/12/genetics-4-blood-cell.jpg)